Alpha-amino acids are the building blocks of proteins. A protein forms via the condensation of amino acids to form a chain of amino acid "residues" linked by peptide bonds. Proteins are defined by their unique sequence of amino acid residues; this sequence is the primary structure of the protein. Just as the letters of the alphabet can be combined to form an almost endless variety of words, amino acids can be linked in varying sequences to form a huge variety of proteins.
Twenty standard amino acids are used by cells in protein biosynthesis, and these are specified by the general genetic code. These twenty amino acids are biosynthesized from other molecules, but organisms differ in which ones they can synthesize and which ones must be provided in their diet. The ones that cannot be synthesized by an organism are called essential amino acids.
Functions in proteins
See also: Primary structure and Posttranslational modification
A polypeptide is a chain of amino acids.
Amino acids are the basic structural building units of proteins. They form short polymer chains called peptides or longer chains either called polypeptides or proteins. The process of such formation from an mRNA template is known as translation which is part of protein biosynthesis. Twenty amino acids are encoded by the standard genetic code and are called proteinogenic or standard amino acids. Other amino acids contained in proteins are usually formed by post-translational modification, which is modification after translation in protein synthesis. These modifications are often essential for the function or regulation of a protein; for example, the carboxylation of glutamate allows for better binding of calcium cations, and the hydroxylation of proline is critical for maintaining connective tissues and responding to oxygen starvation. Such modifications can also determine the localization of the protein, e.g., the addition of long hydrophobic groups can cause a protein to bind to a phospholipid membrane.
Non-protein functions
The twenty standard amino acids are either used to synthesize proteins and other biomolecules, or oxidized to urea and carbon dioxide as a source of energy.[2] The oxidation pathway starts with the removal of the amino group by a transaminase, the amino group is then fed into the urea cycle. The other product of transamidation is a keto acid that enters the citric acid cycle.[3] Glucogenic amino acids can also be converted into glucose, through gluconeogenesis.[4]
Hundreds of types of non-protein amino acids have been found in nature and they have multiple functions in living organisms. Microorganisms and plants can produce uncommon amino acids. In microbes, examples include 2-aminoisobutyric acid and lanthionine, which is a sulfide-bridged alanine dimer. Both these amino acids are both found in peptidic lantibiotics such as alamethicin.[5] While in plants, 1-Aminocyclopropane-1-carboxylic acid is a small disubstituted cyclic amino acid that is a key intermediate in the production of the plant hormone ethylene.[6]
In humans, non-protein amino acids also have biologically-important roles. Glycine, gamma-aminobutyric acid and glutamate are neurotransmitters and many amino acids are used to synthesize other molecules, for example:
Tryptophan is a precursor of the neurotransmitter serotonin
Glycine is a precursor of porphyrins such as heme
Arginine is a precursor of nitric oxide
Carnitine is used in lipid transport within a cell,
Ornithine and S-adenosylmethionine are precursors of polyamines,
Homocysteine is an intermediate in S-adenosylmethionine recycling
Also present are hydroxyproline, hydroxylysine, and sarcosine. The thyroid hormones are also alpha-amino acids.
Some amino acids have even been detected in meteorites, especially in a type known as carbonaceous chondrites.[7] This observation has prompted the suggestion that life may have arrived on earth from an extraterrestrial source.
16 Ocak 2008 Çarşamba
{kind=link}
General Structure
Further information: List of standard amino acids
The general structure of an α-amino acid, with the amino group on the left and the carboxyl group on the right.
In the structure shown to the right, the R represents a side chain specific to each amino acid. The central carbon atom called Cα is a chiral central carbon atom (with the exception of glycine) to which the two termini and the R-group are attached. Amino acids are usually classified by the properties of the side chain into four groups. The side chain can make them behave like a weak acid, a weak base, a hydrophile if they are polar, and hydrophobe if they are nonpolar. The chemical structures of the 20 standard amino acids, along with their chemical properties, are catalogued in the list of standard amino acids.
The phrase "branched-chain amino acids" or BCAA is sometimes used to refer to the amino acids having aliphatic side-chains that are non-linear, these are leucine, isoleucine and valine. Proline is the only proteinogenic amino acid whose side group links to the α-amino group, and thus is also the only proteinogenic amino acid containing a secondary amine at this position. Proline has sometimes been termed an imino acid, but this is not correct in the current nomenclature.[8]
The two optical isomers of alanine.
Isomerism
Most amino acids can exist in either of two optical isomers, called D and L. The L-amino acids represent the vast majority of amino acids found in proteins. D-amino acids are found in some proteins produced by exotic sea-dwelling organisms, such as cone snails.[9] They are also abundant components of the peptidoglycan cell walls of bacteria.[10]
The L and D conventions for amino acid configuration do not refer to the optical activity of the amino acid itself, but rather to the optical activity of the isomer of glyceraldehyde having the same stereochemistry as the amino acid. S-Glyceraldehyde is levorotary, and R-glyceraldehyde is dexterorotary, and so S-amino acids are called L- even if they are not levorotary, and R-amino acids are likewise called D- even if they are not dexterorotary.
There are two exceptions to these general rules of amino acid isomerism. Firstly, glycine, where R = H, no isomerism is possible because the alpha-carbon bears two identical groups (hydrogen). Secondly, in cysteine, the L = S and D = R assignment is reversed to L = R and D = S. Cysteine is structured similarly (with respect to glyceraldehyde) to the other amino acids but the sulfur atom alters the interpretation of the Cahn-Ingold-Prelog priority rule.
{kind=link}
The general structure of an α-amino acid, with the amino group on the left and the carboxyl group on the right.
In the structure shown to the right, the R represents a side chain specific to each amino acid. The central carbon atom called Cα is a chiral central carbon atom (with the exception of glycine) to which the two termini and the R-group are attached. Amino acids are usually classified by the properties of the side chain into four groups. The side chain can make them behave like a weak acid, a weak base, a hydrophile if they are polar, and hydrophobe if they are nonpolar. The chemical structures of the 20 standard amino acids, along with their chemical properties, are catalogued in the list of standard amino acids.
The phrase "branched-chain amino acids" or BCAA is sometimes used to refer to the amino acids having aliphatic side-chains that are non-linear, these are leucine, isoleucine and valine. Proline is the only proteinogenic amino acid whose side group links to the α-amino group, and thus is also the only proteinogenic amino acid containing a secondary amine at this position. Proline has sometimes been termed an imino acid, but this is not correct in the current nomenclature.[8]
{kind=link}
The two optical isomers of alanine.
Isomerism
Most amino acids can exist in either of two optical isomers, called D and L. The L-amino acids represent the vast majority of amino acids found in proteins. D-amino acids are found in some proteins produced by exotic sea-dwelling organisms, such as cone snails.[9] They are also abundant components of the peptidoglycan cell walls of bacteria.[10]
The L and D conventions for amino acid configuration do not refer to the optical activity of the amino acid itself, but rather to the optical activity of the isomer of glyceraldehyde having the same stereochemistry as the amino acid. S-Glyceraldehyde is levorotary, and R-glyceraldehyde is dexterorotary, and so S-amino acids are called L- even if they are not levorotary, and R-amino acids are likewise called D- even if they are not dexterorotary.
There are two exceptions to these general rules of amino acid isomerism. Firstly, glycine, where R = H, no isomerism is possible because the alpha-carbon bears two identical groups (hydrogen). Secondly, in cysteine, the L = S and D = R assignment is reversed to L = R and D = S. Cysteine is structured similarly (with respect to glyceraldehyde) to the other amino acids but the sulfur atom alters the interpretation of the Cahn-Ingold-Prelog priority rule.
Reaction
As amino acids have both a primary amine group and a primary carboxyl group, these chemicals can undergo most of the reactions associated with these functional groups. These include nucleophilic addition, amide bond formation and imine formation for the amine group and esterification, amide bond formation and decarboxylation for the carboxylic acid group. The multiple side chains of amino acids can also undergo chemical reactions. The types of these reactions are determined by the groups on these side chains and are discussed in the articles dealing with each specific type of amino acid.
Peptide bond formation
The condensation of two amino acids to form a peptide bond.
For more details on this topic, see Peptide bond.
As both the amine and carboxylic acid groups of amino acids can react to form amide bonds, one amino acid molecule can react with another and become joined through an amide linkage. This polymerization of amino acids is what creates proteins. This condensation reaction yields the newly formed peptide bond and a molecule of water. In cells, this reaction does not occur directly, instead the amino acid is activated by attachment to a transfer RNA molecule through an ester bond. This aminoacyl-tRNA is produced in an ATP-dependent reaction carried out by an aminoacyl tRNA synthetase.[11] This aminoacyl-tRNA is then a substrate for the ribosome, which catalyzes the attack of the amino group of the elongating protein chain on the ester bond.[12] As a result of this mechanism, all proteins are synthesized starting at their N-terminus and moving towards their C-terminus.
However, not all peptide bonds are formed in this way. In a few cases peptides are synthesized by specific enzymes. For example, the tripeptide glutathione is an essential part of the defenses of cells against oxidative stress. This peptide is synthesized in two steps from free amino acids.[13] In the first step gamma-glutamylcysteine synthetase condenses cysteine and glutamic acid through a peptide bond formed between the side-chain carboxyl of the glutamate (the gamma carbon of this side chain) and the amino group of the cysteine. This dipeptide is then condensed with glycine by glutathione synthetase to form glutathione.[14]
In chemistry, peptides are synthesized by a variety of reactions. One of the most used in solid-phase peptide synthesis, which uses the aromatic oxime derivatives of amino acids as activated units. These are added in sequence onto the growing peptide chain, which is attached to a solid resin support.[15]
Zwitterions
An amino acid, in its (1) normal (unionized) and (2) zwitterionic forms.
As amino acids have both the active groups of an amine and a carboxylic acid they can be considered both acid and base (though their natural pH is usually influenced by the R group). At a certain pH known as the isoelectric point, the amine group gains a positive charge (is protonated) and the acid group a negative charge (is deprotonated). The exact value is specific to each different amino acid. This ion is known as a zwitterion, which comes from the German word Zwitter meaning "hybrid". A zwitterion can be extracted from the solution as a white crystalline structure with a very high melting point, due to its dipolar nature. Near-neutral physiological pH allows most free amino acids to exist as zwitterions.
Peptide bond formation
The condensation of two amino acids to form a peptide bond.
{kind=link}
For more details on this topic, see Peptide bond.
As both the amine and carboxylic acid groups of amino acids can react to form amide bonds, one amino acid molecule can react with another and become joined through an amide linkage. This polymerization of amino acids is what creates proteins. This condensation reaction yields the newly formed peptide bond and a molecule of water. In cells, this reaction does not occur directly, instead the amino acid is activated by attachment to a transfer RNA molecule through an ester bond. This aminoacyl-tRNA is produced in an ATP-dependent reaction carried out by an aminoacyl tRNA synthetase.[11] This aminoacyl-tRNA is then a substrate for the ribosome, which catalyzes the attack of the amino group of the elongating protein chain on the ester bond.[12] As a result of this mechanism, all proteins are synthesized starting at their N-terminus and moving towards their C-terminus.
However, not all peptide bonds are formed in this way. In a few cases peptides are synthesized by specific enzymes. For example, the tripeptide glutathione is an essential part of the defenses of cells against oxidative stress. This peptide is synthesized in two steps from free amino acids.[13] In the first step gamma-glutamylcysteine synthetase condenses cysteine and glutamic acid through a peptide bond formed between the side-chain carboxyl of the glutamate (the gamma carbon of this side chain) and the amino group of the cysteine. This dipeptide is then condensed with glycine by glutathione synthetase to form glutathione.[14]
In chemistry, peptides are synthesized by a variety of reactions. One of the most used in solid-phase peptide synthesis, which uses the aromatic oxime derivatives of amino acids as activated units. These are added in sequence onto the growing peptide chain, which is attached to a solid resin support.[15]
Zwitterions
An amino acid, in its (1) normal (unionized) and (2) zwitterionic forms.
{kind=link}
As amino acids have both the active groups of an amine and a carboxylic acid they can be considered both acid and base (though their natural pH is usually influenced by the R group). At a certain pH known as the isoelectric point, the amine group gains a positive charge (is protonated) and the acid group a negative charge (is deprotonated). The exact value is specific to each different amino acid. This ion is known as a zwitterion, which comes from the German word Zwitter meaning "hybrid". A zwitterion can be extracted from the solution as a white crystalline structure with a very high melting point, due to its dipolar nature. Near-neutral physiological pH allows most free amino acids to exist as zwitterions.
Hydrophilic and hydrophobic amino acids
Depending on the polarity of the side chain, amino acids vary in their hydrophilic or hydrophobic character. These properties are important in protein structure and protein-protein interactions. The importance of the physical properties of the side chains comes from the influence this has on the amino acid residues' interactions with other structures, both within a single protein and between proteins. The distribution of hydrophilic and hydrophobic amino acids determines the tertiary structure of the protein, and their physical location on the outside structure of the proteins influences their quaternary structure. For example, soluble proteins have surfaces rich with polar amino acids like serine and threonine, while integral membrane proteins tend to have outer ring of hydrophobic amino acids that anchors them into the lipid bilayer, and proteins anchored to the membrane have a hydrophobic end that locks into the membrane. Similarly, proteins that have to bind to positively-charged molecules have surfaces rich with negatively charged amino acids like glutamate and aspartate, while proteins binding to negatively-charged molecules have surfaces rich with positively charged chains like lysine and arginine. Recently a new scale of hydrophobicity based on the free energy of hydrophobic association has been proposed.[16]
Hydrophilic and hydrophobic interactions of the proteins do not have to rely only on the sidechains of amino acids themselves. By various posttranslational modifications other chains can be attached to the proteins, forming hydrophobic lipoproteins or hydrophilic glycoproteins.
Hydrophilic and hydrophobic interactions of the proteins do not have to rely only on the sidechains of amino acids themselves. By various posttranslational modifications other chains can be attached to the proteins, forming hydrophobic lipoproteins or hydrophilic glycoproteins.
Nonstandard amino acids
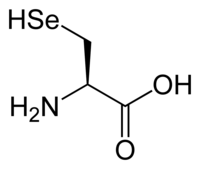
Aside from the twenty standard amino acids, there are a vast number of "non-standard" amino acids. Two of these can be specified by the genetic code, but are rather rare in proteins. Selenocysteine is incorporated into some proteins at a UGA codon, which is normally a stop codon.[18] Pyrrolysine is used by some methanogenic archaea in enzymes that they use to produce methane. It is coded for with the codon UAG.[19]
Examples of nonstandard amino acids that are not found in proteins include lanthionine, 2-aminoisobutyric acid, dehydroalanine and the neurotransmitter gamma-aminobutyric acid. Nonstandard amino acids often occur as intermediates in the metabolic pathways for standard amino acids — for example ornithine and citrulline occur in the urea cycle, part of amino acid catabolism.[20]
Nonstandard amino acids are usually formed through modifications to standard amino acids. For example, homocysteine is formed through the transsulfuration pathway or by the demethylation of methionine via the intermediate metabolite S-adenosyl methionine, [21] while dopamine is synthesized from l-DOPA, and hydroxyproline is made by a posttranslational modification of proline.[22]
Examples of nonstandard amino acids that are not found in proteins include lanthionine, 2-aminoisobutyric acid, dehydroalanine and the neurotransmitter gamma-aminobutyric acid. Nonstandard amino acids often occur as intermediates in the metabolic pathways for standard amino acids — for example ornithine and citrulline occur in the urea cycle, part of amino acid catabolism.[20]
Nonstandard amino acids are usually formed through modifications to standard amino acids. For example, homocysteine is formed through the transsulfuration pathway or by the demethylation of methionine via the intermediate metabolite S-adenosyl methionine, [21] while dopamine is synthesized from l-DOPA, and hydroxyproline is made by a posttranslational modification of proline.[22]
Nutritional İmportance
Further information: Protein in nutrition
Of the 20 standard proteinogenic amino acids, 8 are called essential amino acids because the human body cannot synthesize them from other compounds at the level needed for normal growth, so they must be obtained from food.[23] However, the situation is a little more complicated since cysteine, tyrosine, histidine and arginine are semiessential amino acids in children, because the metabolic pathways that synthesize these amino acids are not fully developed.[24] The amounts required also depend on the age and health of the individual, so it is hard to make general statements about the dietary requirement for some amino acids.
Several common mnemonics have evolved for remembering the ten amino acids often described as essential. PVT TIM HALL ("Private Tim Hall") uses the first letter of each of these amino acids.[27] Another mnemonic that frequently occurs in student practice materials beneath "TV TILL Past Midnight", which just includes the 8 essential amino acids, is "These ten valuable amino acids have long preserved life in man".[28]
Of the 20 standard proteinogenic amino acids, 8 are called essential amino acids because the human body cannot synthesize them from other compounds at the level needed for normal growth, so they must be obtained from food.[23] However, the situation is a little more complicated since cysteine, tyrosine, histidine and arginine are semiessential amino acids in children, because the metabolic pathways that synthesize these amino acids are not fully developed.[24] The amounts required also depend on the age and health of the individual, so it is hard to make general statements about the dietary requirement for some amino acids.
Several common mnemonics have evolved for remembering the ten amino acids often described as essential. PVT TIM HALL ("Private Tim Hall") uses the first letter of each of these amino acids.[27] Another mnemonic that frequently occurs in student practice materials beneath "TV TILL Past Midnight", which just includes the 8 essential amino acids, is "These ten valuable amino acids have long preserved life in man".[28]
See Also
Biochemistry
Beta amino acid
Strecker amino acid synthesis
Glucogenic amino acid
Murchison meteorite, an extraterrestrial source of amino acids.
Table of codons, 3-nucleotide sequences that encode each amino acid
List of standard amino acids (including chemical structures)
Amino acid dating
[edit] References and notes
Beta amino acid
Strecker amino acid synthesis
Glucogenic amino acid
Murchison meteorite, an extraterrestrial source of amino acids.
Table of codons, 3-nucleotide sequences that encode each amino acid
List of standard amino acids (including chemical structures)
Amino acid dating
[edit] References and notes
Reference and Notes
^ Proline is an exception to this general formula. It lacks the NH2 group because of the cyclization of the side chain.
^ Sakami W, Harrington H. "Amino acid metabolism". Annu Rev Biochem 32: 355-98. PMID 14144484.
^ Brosnan J (2000). "Glutamate, at the interface between amino acid and carbohydrate metabolism". J Nutr 130 (4S Suppl): 988S-90S. PMID 10736367.
^ Young V, Ajami A (2001). "Glutamine: the emperor or his clothes?". J Nutr 131 (9 Suppl): 2449S-59S; discussion 2486S-7S. PMID 11533293.
^ Whitmore L, Wallace B (2004). "Analysis of peptaibol sequence composition: implications for in vivo synthesis and channel formation.". Eur Biophys J 33 (3): 233-7. PMID 14534753.
^ Alexander L, Grierson D (2002). "Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening". J Exp Bot 53 (377): 2039-55. PMID 12324528.
^ Llorca J (2004). "Organic matter in meteorites.". Int Microbiol 7 (4): 239-48. PMID 15666244.
^ Claude Liebecq (Ed) Biochemical Nomenclature and Related Documents, 2nd edition, Portland Press, 1992, pages 39-69 ISBN 978-1855780057
^ Pisarewicz K, Mora D, Pflueger F, Fields G, Marí F (2005). "Polypeptide chains containing D-gamma-hydroxyvaline.". J Am Chem Soc 127 (17): 6207-15. PMID 15853325.
^ van Heijenoort J (2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan.". Glycobiology 11 (3): 25R-36R. PMID 11320055.
^ Ibba M, Söll D (2001). "The renaissance of aminoacyl-tRNA synthesis". EMBO Rep 2 (5): 382-7. PMID 11375928.
^ Lengyel P, Söll D (1969). "Mechanism of protein biosynthesis". Bacteriol Rev 33 (2): 264-301. PMID 4896351.
^ Wu G, Fang Y, Yang S, Lupton J, Turner N (2004). "Glutathione metabolism and its implications for health". J Nutr 134 (3): 489-92. PMID 14988435.
^ Meister A (1988). "Glutathione metabolism and its selective modification". J Biol Chem 263 (33): 17205–8. PMID 3053703.
^ Carpino, L. A. (1992) 1-Hydroxy-7-azabenzotriazole. An efficient Peptide Coupling Additive. J. Am. Chem. Soc. 115, 4397-4398.
^ Urry, D. W. (2004). "The change in Gibbs free energy for hydrophobic association - Derivation and evaluation by means of inverse temperature transitions". Chemical Physics Letters 399 (1-3): 177-183.
^ Kyte J & RF Doolittle (1982). "A simple method for displaying the hydropathic character of a protein". J. Mol. Biol. (157): 105-132. PMID 7108955.
^ Driscoll D, Copeland P. "Mechanism and regulation of selenoprotein synthesis.". Annu Rev Nutr 23: 17-40. PMID 12524431.
^ Krzycki J (2005). "The direct genetic encoding of pyrrolysine.". Curr Opin Microbiol 8 (6): 706-12. PMID 16256420.
^ Curis E, Nicolis I, Moinard C, Osowska S, Zerrouk N, Bénazeth S, Cynober L (2005). "Almost all about citrulline in mammals". Amino Acids 29 (3): 177-205. PMID 16082501.
^ Brosnan J, Brosnan M (2006). "The sulfur-containing amino acids: an overview". J Nutr 136 (6 Suppl): 1636S-1640S. PMID 16702333.
^ Kivirikko K, Pihlajaniemi T. "Collagen hydroxylases and the protein disulfide isomerase subunit of prolyl 4-hydroxylases". Adv Enzymol Relat Areas Mol Biol 72: 325-98. PMID 9559057.
^ Young VR (1994). "Adult amino acid requirements: the case for a major revision in current recommendations". J. Nutr. 124 (8 Suppl): 1517S–1523S. PMID 8064412.
^ Imura K, Okada A (1998). "Amino acid metabolism in pediatric patients". Nutrition 14 (1): 143-8. PMID 9437700.
^ Fürst P, Stehle P (2004). "What are the essential elements needed for the determination of amino acid requirements in humans?". J. Nutr. 134 (6 Suppl): 1558S–1565S. PMID 15173430.
^ Reeds PJ (2000). "Dispensable and indispensable amino acids for humans". J. Nutr. 130 (7): 1835S–40S. PMID 10867060.
^ Chapter 39 PVT TIM HALL. Retrieved on 2007-09-29.
^ Memory aids for medical biochemistry. http://mednote.co.kr/Yellownote/BIOCHMNEMON.htm Access date 25 February 2006
^ Sakami W, Harrington H. "Amino acid metabolism". Annu Rev Biochem 32: 355-98. PMID 14144484.
^ Brosnan J (2000). "Glutamate, at the interface between amino acid and carbohydrate metabolism". J Nutr 130 (4S Suppl): 988S-90S. PMID 10736367.
^ Young V, Ajami A (2001). "Glutamine: the emperor or his clothes?". J Nutr 131 (9 Suppl): 2449S-59S; discussion 2486S-7S. PMID 11533293.
^ Whitmore L, Wallace B (2004). "Analysis of peptaibol sequence composition: implications for in vivo synthesis and channel formation.". Eur Biophys J 33 (3): 233-7. PMID 14534753.
^ Alexander L, Grierson D (2002). "Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening". J Exp Bot 53 (377): 2039-55. PMID 12324528.
^ Llorca J (2004). "Organic matter in meteorites.". Int Microbiol 7 (4): 239-48. PMID 15666244.
^ Claude Liebecq (Ed) Biochemical Nomenclature and Related Documents, 2nd edition, Portland Press, 1992, pages 39-69 ISBN 978-1855780057
^ Pisarewicz K, Mora D, Pflueger F, Fields G, Marí F (2005). "Polypeptide chains containing D-gamma-hydroxyvaline.". J Am Chem Soc 127 (17): 6207-15. PMID 15853325.
^ van Heijenoort J (2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan.". Glycobiology 11 (3): 25R-36R. PMID 11320055.
^ Ibba M, Söll D (2001). "The renaissance of aminoacyl-tRNA synthesis". EMBO Rep 2 (5): 382-7. PMID 11375928.
^ Lengyel P, Söll D (1969). "Mechanism of protein biosynthesis". Bacteriol Rev 33 (2): 264-301. PMID 4896351.
^ Wu G, Fang Y, Yang S, Lupton J, Turner N (2004). "Glutathione metabolism and its implications for health". J Nutr 134 (3): 489-92. PMID 14988435.
^ Meister A (1988). "Glutathione metabolism and its selective modification". J Biol Chem 263 (33): 17205–8. PMID 3053703.
^ Carpino, L. A. (1992) 1-Hydroxy-7-azabenzotriazole. An efficient Peptide Coupling Additive. J. Am. Chem. Soc. 115, 4397-4398.
^ Urry, D. W. (2004). "The change in Gibbs free energy for hydrophobic association - Derivation and evaluation by means of inverse temperature transitions". Chemical Physics Letters 399 (1-3): 177-183.
^ Kyte J & RF Doolittle (1982). "A simple method for displaying the hydropathic character of a protein". J. Mol. Biol. (157): 105-132. PMID 7108955.
^ Driscoll D, Copeland P. "Mechanism and regulation of selenoprotein synthesis.". Annu Rev Nutr 23: 17-40. PMID 12524431.
^ Krzycki J (2005). "The direct genetic encoding of pyrrolysine.". Curr Opin Microbiol 8 (6): 706-12. PMID 16256420.
^ Curis E, Nicolis I, Moinard C, Osowska S, Zerrouk N, Bénazeth S, Cynober L (2005). "Almost all about citrulline in mammals". Amino Acids 29 (3): 177-205. PMID 16082501.
^ Brosnan J, Brosnan M (2006). "The sulfur-containing amino acids: an overview". J Nutr 136 (6 Suppl): 1636S-1640S. PMID 16702333.
^ Kivirikko K, Pihlajaniemi T. "Collagen hydroxylases and the protein disulfide isomerase subunit of prolyl 4-hydroxylases". Adv Enzymol Relat Areas Mol Biol 72: 325-98. PMID 9559057.
^ Young VR (1994). "Adult amino acid requirements: the case for a major revision in current recommendations". J. Nutr. 124 (8 Suppl): 1517S–1523S. PMID 8064412.
^ Imura K, Okada A (1998). "Amino acid metabolism in pediatric patients". Nutrition 14 (1): 143-8. PMID 9437700.
^ Fürst P, Stehle P (2004). "What are the essential elements needed for the determination of amino acid requirements in humans?". J. Nutr. 134 (6 Suppl): 1558S–1565S. PMID 15173430.
^ Reeds PJ (2000). "Dispensable and indispensable amino acids for humans". J. Nutr. 130 (7): 1835S–40S. PMID 10867060.
^ Chapter 39 PVT TIM HALL. Retrieved on 2007-09-29.
^ Memory aids for medical biochemistry. http://mednote.co.kr/Yellownote/BIOCHMNEMON.htm Access date 25 February 2006
Kaydol:
Kayıtlar (Atom)